裸子植物のデータベース
裸子植物(Gymnosperms)は、種子が子房に包まれていない植物の総称です。主に木本性の植物で構成され、以下のような特徴を持ちます[ 1 ]:
- 種子が露出している(被子植物のように果実に包まれていない)
- イチイやギンナン、イヌマキなど、被子植物の果実に似た肉質の構造が見られるが、これらは仮種皮や偽果皮と呼ばれる種子の一部。植物学的には裸子植物の種子とその付属構造であり、被子植物の果実とは異なる進化の過程を経て形成されたもの。
- 裸子植物の生殖器官は、花被(がくや花弁)に包まれていない(真の花を持たない)
- 裸子植物の生殖器官は「花」ではなく、「球果」(cone)や「ストロビルス」(strobilus)と呼ばれる
- 受粉は主に風媒
- 木部に仮道管を持つ
- 裸子植物の多くの裸子植物が菌根を形成している
- グネツム類の一部やウェルウィッチアは菌根を形成しない
- 現在知られている裸子植物の中で、明確に草本と分類されるものはない
- 古代植物の遺存種(生き残り)である
裸子植物は約1000種と言われています。被子植物の約35万種と比べるとその0.3 %程でしかありません。
裸子植物には、地球上で最も大きく、最も高い樹木が含まれています。セコイアデンドロン・ギガンテウム(ジャイアントセコイア)Sequoiadendron giganteumは、体積で世界最大の樹木です。一方、セコイア・センペルヴィレンス(コーストレッドウッド)Sequoia sempervirensは、高さで世界最高の樹木であり、100メートルを超える個体も多く存在します。
その他の巨大な裸子植物には、以下のようなものがあります:
- アガチス・アウストラリス(カウリ)Agathis australis:ニュージーランド原産の巨木
- プセウドツガ・メンジージー(ダグラスファー)Pseudotsuga menziesii:北米西部の高木
- ポドカルプス・トタラ(トタラ)Podocarpus totara:ニュージーランドの長寿命の樹木
これらの巨大樹は、森林のバイオマスに大きく寄与し、地球全体のバイオマスの15-20%を占めます。また、生態系において重要な役割を果たし、多くの生物の生息地となっています。炭素固定や気候調節にも大きく貢献しています。ここでは、裸子植物の系統分類と進化について、最新の研究成果を覗いてみます。また、この分野における未解決の問題や今後の研究課題についても考察します。
系統分類の注意点
裸子植物約1000種を系統分類する際に、被子植物と比較して以下のような注意点があります。
- 時間スケールの違い:
裸子植物の進化は被子植物よりもはるかに古く、約3億5000万年前にさかのぼります。一方、被子植物の起源は比較的新しく、約1億4000万年前です。この時間の差は、利用可能な化石記録や分子データの量と質に大きな影響を与えます。 - 多様性と絶滅率の違い:
裸子植物は現在、約1,000種しか存在しませんが、被子植物は35万種以上存在します。裸子植物の多くの系統が絶滅している一方で、被子植物はより多くの系統が現存しています。これにより、被子植物では現存種の遺伝子解析だけでも、より包括的な系統関係の理解ができます。しかしながら、裸子植物の場合、包括的な系統関係を知るうえで絶滅種の遺伝子情報が重要になります。 - 進化速度の違い:
被子植物は裸子植物よりも一般的に進化速度が速いとされています。これは、被子植物がより短い世代時間と、より効率的な繁殖戦略を持っているためです。この速い進化速度により、被子植物の遺伝子には比較的最近の進化の歴史がより明確に刻まれています。 - 分子データの利用可能性:
現存する被子植物の種数が多いため、より多様な分子データが利用可能です。これにより、現存種のデータだけでも、より詳細で信頼性の高い系統樹を構築することが可能になっています。 - 最新の系統分類に用語が追いついていない:
2021年時点で、分子系統学に基づく目-科-属の名前は整理されていません。従来の分類名が使用されます[ 2 ]。
主要なグループと単系統問題
現生の裸子植物は、以下の4つの主要なグループに分類されます:
- ソテツ類 (Cycadophyta)
- イチョウ類 (Ginkgophyta)
- 針葉樹類 (Pinophyta)
- グネツム類 (Gnetophyta)
これらのグループは形態学的にも生態学的にも大きく異なり、それぞれ独自の進化の歴史を持っています。これらのグループをどのように系統分類するか、長年にわたり議論の的となってきました。
主な仮説の変遷は以下の通りです:
- 伝統的分類: 形態学的特徴に基づき、ソテツ類、イチョウ類、針葉樹類、グネツム類を別々のグループとして扱う[ 3 ]。
- アンソフィト仮説: グネツム類を被子植物の姉妹群とする仮説。初期の分子系統解析で支持されたが、現在は否定されている[ 4 ]。
- グネピネ仮説: グネツム類と針葉樹類を姉妹群とする仮説。多くの分子系統解析で支持されてきた[ 5 ]。
- グネチファー仮説: グネツム類、針葉樹類、イチョウ類を一つのクレードとする仮説[ 6 ]。
- 現在の仮説: マツ科とグネツム類を姉妹群とし、他の針葉樹とは別のクレードを形成するとする仮説[ 7 ]。
現在の仮説によれば、裸子植物自体は単系統ながら、マツ科がグネ植物と姉妹群を形成し、他の針葉樹とは別のクレードに属することになり針葉樹類の単系統群が崩れます。
最新の系統樹
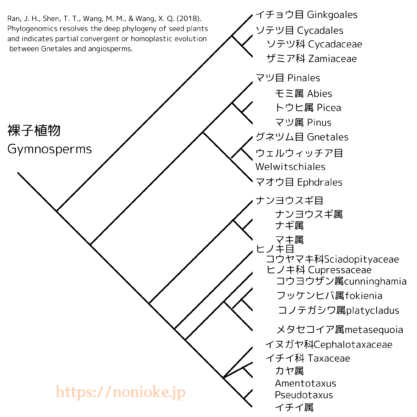
近年の分子系統学的研究により、裸子植物の系統関係についての理解が大きく進展しました。現在最も支持されている系統仮説は、Ran et al. (2018) の研究に基づくものです[ 7 ]。この研究では、大規模なゲノムデータを用いて以下のような系統関係が示されました:
- マツ科 (Pinaceae) とグネツム類 (Gnetales) が姉妹群を形成
- 上記のクレードと他の針葉樹が姉妹群関係
- イチョウとソテツが針葉樹+グネツム類のクレードの姉妹群として分岐
この結果は、裸子植物自体は単系統ながら、針葉樹類は単系統群ではないことを示しています。また、他の遺伝子解析でもこの結果が支持されています[ 8 ][ 9 ][ 10 ]。
裸子植物のデータベース
被子植物のAPGのような国際的な系統分類機関はありませんが、Gymnosperm Database で最新の系統学的知見を反映した情報が提供されています。
裸子植物の目-科-属
分子系統学に基づく目-科-属の名称は、まだ定まっていません。
| 目名 | 科名 | 属 | 種 |
|---|---|---|---|
| ソテツ目 : Cycadales | ソテツ科 : Cycadaceae | 1 | 120 |
| ザミア科 : Zamiaceae | 10 | 246 | |
| イチョウ目 : Ginkgoales | イチョウ科 : Ginkgoaceae | 1 | 1 |
| グネツム目 : Gnetales | ウェルウィッチア科 : Welwitschiaceae | 1 | 1 |
| マオウ科 : Ephedraceae | 1 | 71 | |
| グネツム科 : Gnetaceae | 1 | 40 | |
| マツ目 : Pinales | マツ科 : Pinaceae | 11 | 234 |
| ナンヨウスギ目 : Araucariales | ナンヨウスギ科 : Araucariaceae | 3 | 41 |
| マキ科 : Podocarpaceae | 20 | 174 | |
| ヒノキ目 : Cupressales | コウヤマキ科 : Sciadopityaceae | 1 | 1 |
| ヒノキ科 : Cupressaceae | 28 | 154 | |
| イチイ科 : Taxaceae | 6 | 28 | |
| イヌガヤ科 : Cephalotaxaceae | 3 | 20 |
裸子植物の進化
起源と初期の多様化
裸子植物の起源は古生代デボン紀後期(約3億8500万年前)にさかのぼります。最古の種子植物化石はこの時代のものです。その後、石炭紀から二畳紀にかけて、初期の裸子植物が急速に多様化しました[ 11 ][ 12 ]。
主要な進化イベント:
- 中期ペンシルベニア紀 (約3億1000万年前): 初期の針葉樹、ソテツ、イチョウ類の出現
- ペルム紀 (2億9900万~2億5100万年前): イチョウ目の出現
- 下部ペルム紀 (2億9900万~2億7100万年前): より特徴的なソテツ類の出現
- ペルム紀中期 (2億7100万~2億6000万年前): グネツム類の最初の証拠 (花粉化石)
中生代の繁栄と衰退
裸子植物は中生代に最盛期を迎えました。特に、ジュラ紀から白亜紀前期にかけて、裸子植物は地球上の植生の主要な構成要素となりました[ 11 ][ 12 ]。
主な出来事:
- ジュラ紀: イチョウ類とソテツ類の多様性のピーク
- 白亜紀前期: 針葉樹類の多様化と拡大
- 白亜紀後期: 被子植物の台頭と裸子植物の相対的な衰退
新生代以降の適応と生存戦略
新生代に入ると、被子植物が急速に多様化し、多くの生態的地位を占めるようになりました。この中で、裸子植物は以下のような適応戦略を取ることで生き残りました:[ 13 ][ 14 ][ 15 ]
- 厳しい環境への適応:
- 寒冷地や乾燥地など、被子植物が進出しにくい環境への特化
- 貧栄養土壌での生育能力の獲得
- 長寿命戦略:
- 数百年から数千年生存可能な個体の出現
- 安定した環境での長期的な生存
- 裸子植物には、地球上で最も長寿の生物が含まれています。その中でも特に注目されるのが、Pinus longaeva(ブリストルコーンパイン)で、最古の個体は推定樹齢約5000年とされています。しかし、樹齢の特定は必ずしも容易ではありません。年輪を持つ種では比較的正確な推定が可能ですが、湿潤な気候で生育する種では、木の中心部が腐敗してしまい、正確な年齢を特定できないことがあります。例えば、Thuja plicata(ウェスタンレッドシダー)やTaxus baccata(ヨーロッパイチイ)なども、非常に長寿であると考えられていますが、正確な樹齢は不明です。
- また、クローン繁殖する種では、遺伝的に同一の個体が非常に長期間にわたって存続することがあります。タスマニアのLomatia tasmanicaのクローンは43,600年前のものと推定されています。
- これらの老木は、気候変動や環境の変化に関する貴重な情報源となるだけでなく、生態系の安定性や生物多様性の維持にも重要な役割を果たしています。
- 効率的な水分輸送システム:
- 仮道管による効率的な水分輸送
- 乾燥耐性の向上
- 大量種子生産:
- 風媒による広範囲への種子散布
- 豊凶による捕食圧の回避
これらの戦略により、裸子植物は種数は少ないものの、特定の環境では優占種として存続しています。
各グループの特徴と進化
- ソテツ類 (Cycadophyta)


主な種:
- ソテツ:Cycas
- ボウェニア:Bowenia
- ツノミザミア:Ceratozamia
- ディオーン:Dioon
- エンセファラルトス:Encephalartos
- ウロコヤシ:Lepidozamia
- マクロザミア:Macrozamia
- ミクロシカス:Microcycas
- スタンゲリア:Stangeria
- ザミア:Zamia
特徴:
- パームに似た外観
- 二又分枝する葉脈
- 雌雄異株
進化:
- ペルム紀に出現し、中生代に最盛期を迎えました。
- 現在は主に熱帯・亜熱帯に分布し、約300種が現存しています。
- 多くの種が絶滅の危機に瀕しており、保全の対象となっています。
進化的適応:
- 乾燥に強い葉の構造
- 根に窒素固定細菌を共生させる能力
- 有毒物質の生産による捕食者からの防御
- イチョウ類 (Ginkgophyta)

主な種: Ginkgo
特徴:
- 扇形の特徴的な葉
- 現存種は1種のみ (Ginkgo biloba)
進化:
- ペルム紀に出現し、ジュラ紀に最も多様化しました。
- 白亜紀末の大量絶滅を生き延び、現在は中国の一部地域にのみ自生しています。
進化的適応:
- 落葉性による寒冷気候への適応
- 二又分枝する葉脈による効率的な光合成
- 長寿命と強い環境耐性
- 針葉樹類 (Pinophyta)
特徴:
- 針状または鱗片状の葉
- 球果による種子保護
- 針葉樹類は単系統群ではない
主要な科とその種:
- マツ科 (Pinaceae)
 Picea Pungens Hoopsi: photo by nonioke
Picea Pungens Hoopsi: photo by nonioke- モミ:Abies, カタヤ:Cathaya, ヒマラヤスギ:Cedrus, ケテレーリア:Keteleeria, カラマツ:Larix, チョウセンモミモドキ:Nothotsuga, トウヒ:Picea, マツ:Pinus, コウヨウザン:Pseudolarix, ベイマツ:Pseudotsuga, ツガ:Tsuga
- ヒノキ科 (Cupressaceae)
 Sciadopitys: photo by EMUOK
Sciadopitys: photo by EMUOK- コウヤマキ:Sciadopitys, タスマニアスギ:Athrotaxis, アウストロケドルス:Austrocedrus, カリトリス:Callitris, カリトロプシス:Callitropsis, ネズミサシ:Calocedrus, ヒノキ:Chamaecyparis, スギ:Cryptomeria, コウヨウザン:Cunninghamia, イトスギ:Cupressus, ディセルマ:Diselma, アレルセ:Fitzroya, スイショウ:Glyptostrobus, ヘスペロキパリス:Hesperocyparis, ビャクシン:Juniperus, リボケドルス:Libocedrus, メタセコイア:Metasequoia, ミクロビオタ:Microbiota, パプアケドルス:Papuacedrus, コノテガシワ:Platycladus, セコイア:Sequoia, セコイアデンドロン:Sequoiadendron, タイワニア:Taiwania, ラクウショウ:Taxodium, サンダラック:Tetraclinis, ニオイヒバ:Thuja, アスナロ:Thujopsis, ウィドリントニア:Widdringtonia, キサントキパリス:Xanthocyparis, アメントタクサス:Amentotaxus, オーストロタクサス:Austrotaxus, イヌガヤ:Cephalotaxus, シュードタクサス:Pseudotaxus, イチイ:Taxus, カヤ:Torreya
- ナンヨウスギ科 (Araucariaceae)
 Araucaria: photo by Ann999-DESK
Araucaria: photo by Ann999-DESK- ダンマラ:Agathis, ナンヨウスギ:Araucaria, ウォレミアノビリス:Wollemia
- マキ科 (Podocarpaceae)
 Podocarpus macrophyllus: photo by hiro71
Podocarpus macrophyllus: photo by hiro71- アクモピレ:Acmopyle, アフロカルプス:Afrocarpus, ダクリカルプス:Dacrycarpus, ダクリディウム:Dacrydium, ファルカティフォリウム:Falcatifolium, ハロカルプス:Halocarpus, ラガロストロボス:Lagarostrobos, レピドタムヌス:Lepidothamnus, マノアオ:Manoao, ミクロカクリス:Microcachrys, ナギ:Nageia, パラシタクサス:Parasitaxus, ペクチノピティス:Pectinopitys, フェロスファエラ:Pherosphaera, マキバブシ:Phyllocladus, マキ:Podocarpus, プルムノピティス:Prumnopitys, レトロフィルム:Retrophyllum, サクセゴタエア:Saxegothaea, スンダカルプス:Sundacarpus
進化:
- 石炭紀後期に出現し、中生代に急速に多様化しました。
- 現在も温帯から寒帯の森林生態系で重要な役割を果たしています。
進化的適応:
- 仮道管による効率的な水分輸送
- 樹脂の生産による病害虫への防御
- 寒冷・乾燥環境への適応
- グネツム類 (Gnetophyta)



特徴:
- 被子植物に似た特徴を持つ (例: 導管の存在)
- 3つの属 (グネツム属:Gnetum, マオウ属:Ephedra, ウェルウィッチア属:Welwitschia) からなる
進化:
- ペルム紀に出現したとされますが、大型化石の記録は白亜紀以降です。
- 被子植物との収斂進化が見られます。
進化的適応:
- Ephedra: 乾燥環境への適応
- Welwitschia: 極度の乾燥環境での生存戦略
- Gnetum: 熱帯雨林環境への適応
裸子植物の生態学的重要性
生態系における役割
裸子植物、特に針葉樹類は、以下のような重要な生態学的役割を果たしています:
- 炭素固定:
- 大型で長寿命の樹木として、大量の炭素を固定
- 北方林や山岳地帯での主要な一次生産者
- 生息地の提供:
- 多くの動物種の生息地や餌資源となる
- 特に針葉樹林は固有の生態系を形成
- 水循環の調節:
- 森林による水分保持と蒸発散の調節
- 山岳地帯での雪解け水の調節
- 土壌保全:
- 根系による土壌の安定化
- 有機物の供給による土壌形成
人間社会との関わり
裸子植物は人間社会と深い関わりを持っています:
- 木材資源:
- 建築材、家具、紙パルプなどの原料
- 特に針葉樹は主要な木材資源
- 食料:
- ソテツの種子やイチョウの実など、一部は食用
- マツの実は重要な食料資源
- 薬用:
- イチョウ葉エキスなど、伝統医療や現代医薬品の原料
- エフェドラ属植物からのエフェドリン抽出
- 観賞用:
- 庭園や公園での利用
- 盆栽や生け花の材料
- 文化的・宗教的意義:
- 多くの文化で神聖視される樹木 (例: レバノン杉)
- 長寿の象徴としての意味合い
- 日本におけるコウヤマキ
- コウヤマキ(Sciadopitys verticillata)は、日本固有の裸子植物で、非常に興味深い進化的歴史を持っています。コウヤマキ科(Sciadopityaceae)に属する唯一の現存種であり、「生きた化石」とも呼ばれています。
- コウヤマキの特徴:
- 常緑の針葉樹で、高さ30メートルに達することがあります。
- 特徴的な傘状の枝ぶりから、「傘松」とも呼ばれます。
- 葉は実際には短枝が変形したもので、「葉状枝」と呼ばれます。
- 日本での分布と文化的意義:
- 本州、四国、九州の山地に自生しています。
- 特に和歌山県の高野山周辺に多く生育することから、「高野槇(こうやまき)」という名前がつきました。
- 古くから神社仏閣の建築材として珍重されてきました。
- 耐久性が高く、防腐効果もあるため、風呂桶や建具などにも使用されてきました。
- 保全状況:
- 天然林の減少や過剰な伐採により、自然個体群の多くが減少しています。
- 現在は、植林や保護活動が行われていますが、成長が遅いため、回復には時間がかかります。
- コウヤマキは、日本の森林生態系と文化の両面で重要な役割を果たしており、その保全は生物多様性の維持と文化遺産の保護の観点から重要です。
保全と課題
- 絶滅の危機
多くの裸子植物種が絶滅の危機に瀕しています:
- IUCN (2020) によると、針葉樹の約34%、ソテツ類の約63%が絶滅危惧種に分類されています[ 16 ]。
- 特にソテツ類は、生息地の破壊や違法採取により深刻な危機に直面しています。
主な脅威:
- 生息地の破壊と分断
- 気候変動の影響
- 過剰採取 (木材、観賞用、薬用)
- 外来種の侵入
- 病害虫の蔓延
- 保全への取り組み
裸子植物の保全のために、以下のような取り組みが行われています:[ 17 ]
- 生息地の保護:
- 保護区の設定
- 生態系管理の実施
- 種の保全:
- 絶滅危惧種の個体数モニタリング
- 植物園や種子バンクでの系統保存
- 持続可能な利用:
- 森林認証制度の導入
- 代替資源の開発
- 研究と教育:
- 生態学的研究の推進
- 一般市民への啓発活動
- 国際協力:
- CITES (ワシントン条約) による取引規制
- 国際的な保全プログラムの実施
今後の課題
裸子植物の系統と進化に関する理解は大きく進展しましたが、まだ多くの課題が残されています:[ 18 ]
- 深い系統関係の解明:
- 古い時代の化石データと分子データの統合
- 絶滅した系統の位置づけの明確化
- 進化メカニズムの解明:
- ゲノム進化と形態進化の関連性の解明
- 適応放散のプロセスの解明
- 生態学的役割の詳細な理解:
- 気候変動下での裸子植物の応答予測
- 生態系サービスの定量的評価
- 保全生物学的研究:
- 絶滅危惧種の遺伝的多様性の評価
- 効果的な保全戦略の開発
- 古環境の復元:
- 裸子植物の化石記録を用いた過去の気候・環境の推定
- 大量絶滅イベントの影響と回復プロセスの解明
- 共生関係の研究:
- 菌根菌との相互作用の進化
- 昆虫など他の生物との共進化プロセスの解明
- ゲノム編集技術の応用:
- 絶滅危惧種の保全への応用可能性
- 有用形質の改良と新品種開発
まとめ
裸子植物は、地球上の植物相の重要な構成要素であり、その進化の歴史は植物の多様化プロセスを理解する上で極めて重要です。近年の分子系統学的研究により、裸子植物の系統関係についての理解が大きく進展しましたが、まだ多くの謎が残されています。
裸子植物は、被子植物の台頭後も独自の適応戦略を発達させ、特定の環境で重要な生態学的役割を果たし続けています。しかし、多くの種が人間活動や環境変化の影響を受け、絶滅の危機に瀕しています。
今後の研究課題としては、より古い時代の化石データと最新の分子データを統合し、裸子植物の深い系統関係を解明することが挙げられます。また、ゲノムレベルでの進化メカニズムの解明や、気候変動下での裸子植物の応答予測など、多岐にわたる研究が必要とされています。
裸子植物の保全は、生物多様性の維持と生態系機能の保護のために極めて重要です。科学的知見に基づいた効果的な保全戦略の開発と実施が求められています。
最後に、裸子植物の研究は、植物進化の全体像を理解する上で不可欠です。被子植物との比較研究を通じて、種子植物全体の進化プロセスや適応メカニズムについての理解が深まることが期待されます。
用語解説
- 単系統群(クレード): 共通の祖先から派生した生物群
- 姉妹群: 系統樹上で最も近縁な関係にある2つのグループ
- 収斂進化: 異なる系統の生物が、類似した環境に適応した結果、似た形質を獲得すること
- 仮道管: 針葉樹の木部にある水分輸送に特化した細長い細胞
- 菌根: 植物の根と菌類が形成する共生関係
- IUCN: 国際自然保護連合(絶滅危惧種のレッドリストを作成している国際機関)
- CITES: 絶滅のおそれのある野生動植物の種の国際取引に関する条約(ワシントン条約)
参考文献
- The Gymnosperm Database
- Kew Tree of Life Explorer
- An Introduction to Plant Taxonomy
- Is the anthophyte hypothesis alive and well? New evidence from the reproductive structures of Bennettitales
- Seed plant phylogeny: Gnetophytes are derived conifers and a sister group to Pinaceae
- Gnetifer hypothesis
- Ran, J. H., Shen, T. T., Wang, M. M., & Wang, X. Q. (2018). Phylogenomics resolves the deep phylogeny of seed plants and indicates partial convergent or homoplastic evolution between Gnetales and angiosperms. Proceedings of the Royal Society B: Biological Sciences, 285(1881), 20181012.
- One Thousand Plant Transcriptomes Initiative. (2019). One thousand plant transcriptomes and the phylogenomics of green plants. Nature, 574(7780), 679-685.
- Leslie, A. B., Beaulieu, J. M., Crane, P. R., & Donoghue, M. J. (2013). Explaining the distribution of breeding and dispersal syndromes in conifers. Proceedings of the Royal Society B: Biological Sciences, 280(1770), 20131812.
- Wang, X. Q., & Ran, J. H. (2014). Evolution and biogeography of gymnosperms. Molecular Phylogenetics and Evolution, 75, 24-40.
- Condamine, F. L., Silvestro, D., Koppelhus, E. B., & Antonelli, A. (2020). The rise of angiosperms pushed conifers to decline during global cooling. Proceedings of the National Academy of Sciences, 117(46), 28867-28875.
- Crisp, M. D., & Cook, L. G. (2011). Cenozoic extinctions account for the low diversity of extant gymnosperms compared with angiosperms. New Phytologist, 192(4), 997-1009.
- Ecology and evolution of pine life histories
- The tortoise and the hare: ecology of angiosperm dominance and gymnosperm persistence
- Conifer Reproductive Biology
- IUCN. (2020). The IUCN Red List of Threatened Species. Version 2020-3. https://www.iucnredlist.org
- Farjon, A., & Filer, D. (2013). An atlas of the world’s conifers: an analysis of their distribution, biogeography, diversity and conservation status. Brill.
- Mathews, S. (2009). Phylogenetic relationships among seed plants: persistent questions and the limits of molecular data. American Journal of Botany, 96(1), 228-236.
